| 59.4% |  | United States |
| 8.7% |  | United Kingdom |
| 5% |  | Canada |
| 4.1% |  | Australia |
| 3.5% |  | Philippines |
| 2.6% |  | Netherlands |
| 2.4% |  | India |
| 1.6% |  | Germany |
| 1% |  | France |
| 0.7% |  | Poland |
| Today: | 179 |
| Yesterday: | 251 |
| This Week: | 179 |
| Last Week: | 2221 |
| This Month: | 4767 |
| Last Month: | 6796 |
| Total: | 129366 |
3. Psychological and Cognitive Effects of Cannabis
 |
|  |
| 
| Books - Cannabis and Man |
Drug Abuse
3. Psychological and Cognitive Effects of Cannabis
Jared R. Tinklenberg and Charles F. Darky, Department of Psychiatry, Stanford University School of Medicine.
1. INTRODUCTION
Descriptions of the psychological effects of cannabis have been found in Eastern documents and archaeological artifacts dating back to before the time of Christ. Throughout the subsequent centuries writers from various parts of the world have described, at times with remarkable clarity, the subjective and behavioral changes induced by this drug. In recent years, with the application of sophisticated experimental methods, especially the precise quantification of g-tetrahydrocannabinols, the chemicals in cannabis that exert most of the psychoactive effects, our understanding of the drug's psychological and cognitive effects has increased. This paper provides an overview of the present state of knowledge, describes this information in terms of conceptual model, and suggests directions for future cannabis research.
2. BASIC CONSIDERATIONS
The effects of cannabis on psychological and cognitive processes are influenced by a number of variables. Since these variables have been discussed in most recent reviews on cannabis, only a brief summary will be presented here.
Recent experiments have demonstrated that when reliable quantities of smoked cannabis or tetrahydrocannabinol (THC) are delivered to subjects reproducible dose-dependent and time-action effects occur with many psychological and cognitive tasks (Casswell and Marks, 1973; Isbell et al., 1967; Kiplinger et al., 1971; Lemberger et al., 1970; Manno et al., 1971; Renault et al., 1971) . As with most psychoactive drugs, larger doses of cannabis produce greater and more prolonged psychological effects. In addition to dose, the route of administration-- smoked, oral, or injected--also influences the effects of cannabis (Hollister, 1971). With smoking or injection, the onset of psychological effects is almost immediate'and the total duration of action, depending upon dosage, is usually less than 3 to 4 hours. Drugs effects from ingested THC are qualitatively similar but differ quantitatively as a result of individual variations in intestinal absorption, the material in which the THC is dissolved, and the phase of the time-action curve (Perez-Reyes et al., 1973). The peak intensity of ingested cannabis is usually only 1/3 or 1/2 of an equivalent smoked or injected dose.
The individual's previous pattern of cannabis use is another important variable that influences the psychological and cognitive effects of the drug. With repetitive use, tolerance appears to develop to certain of the behavioral effects, that is, some performance deficits with acute intoxication seen in occasional users not demonstrated in regular users (Meyer et al., 1971; Weil et al., 1968). Although early observations indicated the possibility of increased pharmacological sensitivity to the subjective effects of cannabis with repeated use--so-called 'reverse tolerance'--subsequent studies have provided strong evidence of the development of tolerance to the psychological and cognitive as well as physiological effects of cannabis (Hollister, 1974). The enhanced sensitivity to subjective effects reported by chronic users probably results from learned responses rather than from any direct drug effect (Jones, 1974).
The subjective and behavioral effects of cannabis are significantly influenced by 'set'--the attitudes and expectations of the individual user derived from mood, personality, and past drug experiences, and 'setting' the physical and social environment in which drug use takes place. These non-pharmacological variables are especially important at low doses: however, at higher doses pharmacological properties become progressively more dominant. These basic considerations should be remembered in reviewing the psychological and cognitive effects of cannabis described in the following sections.
3. SUBJECTIVE EFFECTS AND EXPERIMENTAL EVIDENCE
The subjective and behavioral effects of cannabis depend heavily on the basic factors just discussed and hence vary considerably among individuals. With low dose intoxication, the most commonly reported subjective effects are initial euphoria, increased hilarity, mild restlessness, exhilaration, and later a dreamy, carefree state of relaxation. Although these pleasant effects are frequently reported, some users experience various degrees of anxiety and other dysphoric feelings.
Altered time sense is one of the most consistent and marked perceptual effects of cannabis. Even with low doses, users almost invariably report that there is an acceleration of subjective time so that geophysical time seems to pass slowly. This effect, which may be interpreted as a speeding up of the hypothetical 'internal clock' so that 'inner time' is fast compared to geophysical time, has been extensively documented with experimental evidence (Canadian Commission, 1972; Cappell et al., 1972; Clark et al., 1970; Hollister and Gillespie, 1970; Hollister and Tinklenberg, 1973; Karniol and Carlini, 1973; Tinklenberg et al., 1972).
Perceptual experience arising from stimulation of sensory receptors also seems altered during cannabis intoxication. Enhanced tactile, olfactory, and gustatory perceptions are often reported. These effects may coalesce to produce the increased hunger, or greater interest in food, often reported during cannabis intoxication. Increased appreciation of the subtle nuances of sound and visual patterns are commonly noted and dose-related increases in subjective reports of imagery have been reported, including increased vividness, clarity, colour, movement, depth, and complexity (Canadian Commission, 1972). Despite these persistent subjective reports of cannabis-induced enhancement of sensory-perceptual processes, objective measures have demonstrated either no change or slight decrements in perceptual functioning (Caldwell et al., 1969, 1970; Moskowitz et al, 1972; Roth et al., 1973). In other words, there is no firm evidence of changes in peripheral sensory thresholds or actual enhancement of perceptual acuity; instead, any increases in perceptual awareness seem to occur at a subsequent stage of processing.
In addition to these subjective effects which entail primarily alterations in the basic sensory modalities, cannabis induces a wide variety of other subjective changes (Tart, 1970). New meanings are found in commonplace events; prosaic ideas seem original. Although many users report that their thinking does not follow logical patterns, they nevertheless feel their reasoning is intuitively correct. Thinking in abstract symbols is reduced while visual imagery is often increased and many users report an increase in spontaneous visual imagery while reading. With higher doses of cannabis, these subjective effects are intensified. The individual may experience rapidly altering emotions, fragmentation of perceptions, an altered sense of self-identity and feelings of enhanced insight. Introspection and social withdrawal are often marked. Synesthesias, the imagery of one sense mode aroused by stimulation of another sense mode, have been reported; for example, the individual may experience colours in response to an auditory stimulus. Dream-like fantasies and vivid hallucinations may occur, usually in a discontinuous or undulating pattern. Laboratory experiments with high doses of THC have produced perceptual phenomena very similar to those induced by LSD including distortions of body image, extreme perceptual aberrations, and a dissolution of personal identity (Isbell et al., 1967; Melges et al., 1970a; Melges et al., 1974; Perez-Reyes et al., 1973).
Experienced users usually describe these effects as pleasant, especially when the factors of set and setting are non-threatening and conducive to a pleasant experience. However, the same cannabis effects may also be interpreted as unpleasant, particularly by neophytes who may fail to identify the subjective and behavioral alterations as transient. Severe anxiety and panic may result when inexperienced users interpret the drug state as signs of permanent psychosis or impending death.
4. CANNABIS AND PSYCHOMOTOR TASKS
Subjective reports by cannabis users indicate that many feel as if their ability to drive vehicles or perform other complex motor tasks is reduced, but that they are usually able to compensate for any impairment (Hochman and Brill, 1971). Laboratory studies have confirmed that performances on a wide range of psychomotor tasks are impaired during cannabis intoxication. Cannabis has induced deficits in tapping speed, handwriting, simple and complex reaction time, pursuit motor and tracking tasks, continuous performance tests, and driver simulation studies (Clark and Nakashima, 1968; Clark et al., 1970); Dornbush et al., 1971; Galanter et al., 1972; Hollister and Gillespie, 1970; Kiplinger et al., 1971; Manno et al., 1970; Melges et al., 1970b; Mendelson and Meyer, 1972; Meyer et al., 1971; Rafaelson et al., 1973; Roth et al., 1973; Volavka et al., 1971; Weil and Zinberg, 1969; Weil et al., 1968). Previous use of cannabis may also significantly influence performances on psychomotor tasks. At low doses, less experienced users generally show greater impairment on psychomotor tasks than do experienced users (Clark and Nakashima, 1968; Jones, 1971; Jones and Stone, 1970; Mendelson and Meyer, 1972; Meyer et al., 1971; Weil and Zinberg, 1969). How much this phenomenon represents the ability of experienced users to volitionally compensate for drug effects and how much it represents other processes, such as the development of tolerance at a pharmacological level, is difficult to determine.
5. MEMORY - COGNITIVE EFFECTS
One of the most commonly reported effects of cannabis intoxication is difficulty in performing tasks in which information recently stored in memory must be accurately retrieved. In 1932 Beringer described cannabis-induced 'fragmentation of thought processes.. .disturbances of memory.. .interruptions in the stream of thought' (Ames, 1958). Bromberg's classic description of cannabis effects noted that ideas flowed so quickly that they failed to become fixed in memory (1934). Similarly the early investigations of the Chopras and others placed special emphasis on alterations in memory and cognitive processes (Chopra and Chopra, 1939). More recent surveys of cannabis effects have confirmed and elaborated these early clinical observations. Many of Tart's subjects reported that their memory span was shortened so that they forgot what a conversation was about before it had ended (1970). Since concepts often slipped away before they could be grasped, thinking lost clarity.
Directed in part by these consistent reports from cannabis users that the drug often interferes with the ability to follow through logical sequences of thought, to engage in conversation, and to recall recent events, researchers have investigated cannabis effects on a variety of memory tasks. The subjective reports and clinical observations were verified in the laboratory; the performance of cannabis subjects was impaired for tasks requiring remembering of simple stimuli for short periods of time such'as digit span (Melges et al., 1970b; Tinklenberg et al., 1970), digit code (Clark and Nakashima, 1968; Clark et al., 1970; Jones and Stone, 1970), and memory scan (Darley et al., 1973a), as well as tasks in which more complex stimuli had to be processed and retained in memory such as Babcock Story Recall (Drew et al., 1972; Miller et al., 1972), picture recognition and Benton Sentence Repetition (Klonoff et al., 1973).
Since basic memory processes are fundamental to a wide range of cognitive functions, a considerable amount of research has been focused on gathering data about cannabis induced changes in memory processes. These
data now require organization into a conceptual framework. Recently, results from several studies utilizing free-recall memory tasks have been interpreted in terms of a model of human memory which allows the effect of cannabis on memory to be more explicitly identified (Atkinson et al., 1973). The structural components of this model are a very short-term sensory register, a limited-capacity short-term store (STS) or working memory, and a large, permanent Long-term store (LTS). (See Figure 3.1) Stimuli from the environment are received by the sensory receptors and held in the sensory register in an unprocessed form for a period of milliseconds (Sperling, 1960). Each stimulus must then be analyzed so that its internal encoded representation, which is retained in LTS, may be located and activated. The portion of LTS which contains the encoded representations of stimuli is called the conceptual store (CS). From the conceptual store of LTS the encoded stimulus representation is entered into working memory (STS). At this point the individual develops conscious awareness of the stimulus. While the stimulus resides in STS it may be operated upon by control processes such as rehearsal (rote repetition) or the formation of mnemonics or visual images. These rehearsal processes place information regarding the stimulus' occurrence as an environmental event into another portion of LTS, the event-knowledge store (EKS). Thus, the EKS of the long term store acquires and maintains information about the context in which a particular item or event occurred. In contrast, the conceptual store (CS) of LTS contains information about primary characteristics and meaning of stimuli.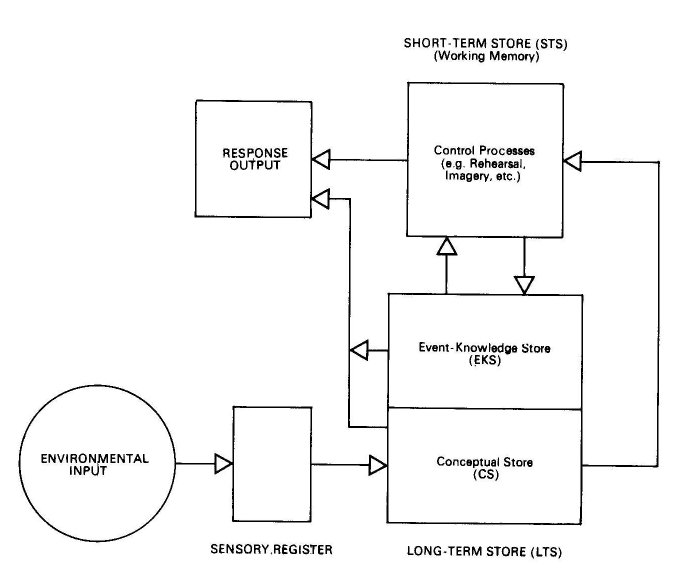
Figure 3.1. Information flow through the memory system begins with the processing of environmental input in the sensory register. The stimulus is analysed and its encoded representation in the conceptual store (CS), a partition of long-term store (LTS), is activated. The encoded representation is then sent to short-term store (STS) where it receives additional processing by means of control processes. Control processes cause information to be copied into the event-knowledge store (EKS), a second partition of LTS. From EKS, information about the context in which the stimulus occurred may be retrieved and entered into STS. Depending upon task requirements, a response may be initiated at any stage of processing, utilizing information retrieved from either CS, STS or EKS.
The longer an event is actively processed in STS, the more information about that event is transferred to EKS. However, when the control processes of STS do not operate upon an encoded stimulus, the stimulus is lost from STS within about 30 seconds. An encoded stimulus may reside in both STS and the EKS portion of LTS simultaneously; information about the stimulus may pass from the EKS to STS as well as vice versa.
In free recall tasks a subject is presented a series of items such as words which he is to recall in any order after the list has been presented. According to the above model, the items (words) are sequentially received by
the sensory receptors, briefly held in the sensory register, encoded in the CS, entered into STS, and transferred to the EKS portion of the LTS by means of control processes. Only a limited number of items can be actively processed in STS, so after the fourth or fifth item has been presented, the arrival of new items causes old items to be dropped from STS. At the time of recall, the retrieval of list items may be from STS, EKS, or both depending on where in the list the item was presented and how soon after list presentation testing occurs. With an immediate free recall test, terminal list items are retrieved mainly from STS and earlier items largely from LTS. When testing is delayed, all items are usually retrieved from LTS (Craik, 1970; Glanzer and Cunitz, 1966).
Cannabis intoxication does not impair the delayed free recall of information which had been stored in LTS in a non-cannabis state (Abel, 1971; Darley et al., 1973b; Dornbush, 1974). In other words, cannabis does not appreciably reduce retrieval of items which have been adequately stored in LTS. In fact, information initially presented to subjects during cannabis intoxication is most efficiently recalled when subjects are again under the influence of cannabis ()arley et al., 1974; Eich et al., 1974). These results indicate that cannabis induces asymetrical state-dependency for retrieval of information with free recall procedures, i.e. information presented in the drug state is more efficiently recalled during drug intoxication than during non-intoxication, but information initially stored during non-drug conditions is recalled equally well in either a drug or non-drug state. However, the influence of cannabis on retrieval processes seems to vary according to the type of memory task utilized since retrieval for paired-associate learning is also asymetrically state-dependent for cannabis (Rickles et al., 1973), while performance on a recognition memory task does not show any type of state dependency (Darley et al., 1974).
Although cannabis does not impair retrieval on delayed recall tests, cannabis does induce significant deficits on immediate recall testing (Abel, 1971; Darley, et al., 1973b; Pearl et al., 1973). Since retrieval is not impaired by cannabis, this finding indicates that the drug interferes with some phase of memory storage. (Dornbush, 1974). Using the model presented earlier, this phase can be identified. Since terminal list items are recalled equally well by cannabis and placebo subjects, neither storage into nor retrieval from STS is appreciably impaired by the drug. Instead, the cannabis induced immediate recall deficits involve non-terminal list items which can be recalled only if they have been transferred from STS to LTS. Thus, the deleterious effect of cannabis on memory storage must be in the phase where information is transferred from STS to LTS.
Since transfer to LTS is controlled by active processes such as rehearsal, a likely explanation for the cannabis effect on memory storage is that drug subjects engage in less rehearsal and other control processes, perhaps due to external and/or internal distractions (Abel, 1971). However, the storage deficit persists when overt fixed-rehearsal procedures are used to equalize the degree to which cannabis and placebo subjects overtly process list items ()arley et al., 1974). Thus, even when cannabis subjects rehearse list items as often as placebo subjects, they store less information about those items.We have suggested that drug subjects may attend to list items only enough to produce the required minimum of rehearsals, while placebo subjects in addition to performing the required overt rehearsals, covertly processed the items for later recall. (Darley and Tinklenberg, 1974). An alternative explanation is that once rehearsal of an item is terminated cannabis causes information which would be transferred to LTS to be lost from STS more quickly. This possibility receives support from a study by Dornbush et al., (1971) which showed that unrehearsed items are lost from STS more quickly during cannabis intoxication (See Darley and Tinklenberg, 1974, for further details on this issue).
Although, as indicated above, certain acute, short-term psychological and cognitive effects of cannabis have been clearly established, little is known about the effects of repetitive, long-term cannabis consumption. Our present paucity of knowledge about this important topic stems directly from the lack of adequate studies. Hopefully, investigations currently being conducted will remedy this situation.
6. FUTURE OF PSYCHOLOGICAL AND COGNITIVE RESEARCH WITH CANNABIS
In considering future avenues of research it is often useful to organize what is presently known into a conceptual sketch or model which lends itself to further experimental manipulation. In attempting this with cannabis, we consider five areas of evidence about the effects of cannabis on human psychological and cognitive processes to be especially salient.
1. Cannabis does not markedly alter the earliest (sensory-perceptual) phases of information processing, at least at peripheral levels.
2. However, cannabis often alters the individual's subjective appreciation or interpretation of many sensory-perceptual experiences.
3. Cannabis markedly changes many components of time sense, especially inducing enhanced subjective tempo so that the duration of events seems to be longer than usual.
4. Cannabis does not significantly impair the retrieval of information which has been adequately stored in LTS .
5. However, cannabis does seem to impair the initial storage of information in LTS, possibly by making control processes such as rehearsal less efficient.
We propose that cannabis may alter the manner in which encoded information is entered into STS thereby causing the 5 salient effects noted above. To develop our proposal we utilize the memory model presented earlier as well as other recent models which describe how information in LTS is encoded, stored, and retrieved (Atkinson et al., 1973; Melton and Martin, 1972; Norman, 1970; Tulving and Donaldson, 1972). When a stimulus, such as a word, makes contact with sensory receptors it is analyzed initially in terms of its perceptual characteristics. As mentioned previously, this analysis allows the internal encoded representation of the stimulus to be located in the conceptual store (CS) of LTS and activated. The internal representation may have a number of codes which specify the value of the stimulus on a set of dimensions or attributes (Norman and Rumelhart, 1970; Underwood, 1969). For words these attributes may be perceptual, syntactic or semantic properties. In most instances the requirements of the subject's task determine which properties of the stimulus are important and which may be ignored. Therefore, only a subset of the codes which fully describe a stimulus may be placed into STS (working memory) from LTS and thus enter the individual's conscious awareness. For example, if a subject is shown a picture of a ball and required to report what he saw after a short delay, he may enter into STS only the name of the object, i.e. 'ball' and ignore other characteristics such as size, shape and color. The way in which the stimulus is encoded determines to a great extent the subjective perception of the stimulus and the way in which it is processed by the memory stores. Since items with common characteristics are thought to share some common codes in LTS, the activation of a set of codes for a stimulus is likely to activate codes representing related items. The entry into STS of items associated to the presented stimulus has been termed an 'implicit associative response' (JAR) (Bousfield et al., 1958; Underwood, 1965). For example, if the word 'dog' were presented on a list, the word 'cat' might enter STS (working memory) as an IAR since the two items have some common perceptual and semantic features.
We propose that cannabis causes more codes to be entered into STS for each new item presented. For example, the presentation of the word 'apple' to a
subject under the influence of cannabis might cause a variety of perceptual and semantic information to enter STS such as 'it's a fruit called an apple which is round, red, shiny,
and sweet tasting', whereas for a subject who has not taken cannabis only the name of the stimulus might enter STS (working memory). The more codes activated for a particular stimulus, the more likely it is that codes for related items will also be activated and enter STS. This model may be useful in explaining several different psychological effects of cannabis.
First, it is consistent with a limited amount of directly applicable experimental evidence which indicates that during cannabis intoxication there is an increase in intrusion errors, i.e., the subject is likely to erroneously recall information which was not contained in the initial learning task (Abel, 1971; Darley and Tinklenberg, 1974; Dornbush, 1974; Miller et al., 1972). Such intrusions probably occur because IARs are made at the time of stimulus presentation (Ainisfield and Knapp, 1968). The greater number of IARs which result from the increased range of encoding under cannabis would therefore account for such errors of intrusion.
Second, the model is consistent with most experimental cannabis work which focuses on memory processes. Cannabis-induced storage deficits arise because each newly-presented stimulus expends more of the limited capacity of STS. During cannabis intoxication the entry into STS of a larger number of codes for each stimulus and related non-list items causes codes from earlier list items to be dropped more quickly from STS than during the
non-drug state. Many of the entered codes may be irrelevant for the task requirements; the time spent processing them reduces the time available for processing task-relevant codes. Since the processing applied to an item's relevant codes determines how much information about that item is transferred to LTS, less transfer occurs in the drug state. In addition, since cannabis causes fewer recall items to be represented in STS at any one time, complex control processes which allow list items to be associated with one another become less efficient. Again, the result is less information available in LTS about the occurrence of an item and therefore poorer immediate recall performance for items which must be retrieved from LTS. The model is also consistent with data indicating that cannabis does not impair retrieval of already stored information; if anything, the proposed cannabis effect might enhance retrieval since particular encodings of information previously inaccessible might be located during cannabis intoxication. No objective evidence for enhanced retrieval with cannabis exists at the present time, but appropriate measures may not have been applied.
Third, the cannabis induced alterations in time sense, especially the acceleration of subjective tempo would be explicable with this model. The increased flow of codes into STS would induce the greater frequency of inner events which has been theoretically associated with an acceleration of subjective tempo (Ornstein, 1969; Tinklenberg et al., 1972). In other words, the greater number of items impinging on STS (conscious awareness) per unit of geophysical time would explain the frequently reported effects of cannabis in which geophysical time seems to go more slowly than usual.
Fourth, the proposed model would be compatible with the cannabis-induced changes in sensory-perceptual processes where there is often enhancement of subjective awareness of appreciation in the absence of increased peripheral sensory acuity. The increased flow to STS of usually inaccessible codes for a stimulus and related items might subjectively enhance the richness and uniqueness of sensory events. The synesthetic experiences may also be explicable since under marijuana perceptual codes related to different sensory systems (eg. a visual code and an auditory code) may enter STS together.
Fifth, since conscious thought processes involve interrelating and manipulating concepts, having available to consciousness a wider range of properties for any particular concept and related concepts might also explain the feelings of increased insight and creativity which occur during cannabis intoxication. Intoxicated subjects may be able to connect usually unrelated concepts or perceive new aspects of objects or events.
One prediction which arises from this model is that difficult tasks in which information must be extensively processed in STS should be affected by cannabis while well-learned, automatic tasks in which information may flow directly from LTS to the response apparatus will show little effect. Also, deficits due to an influx of irrelevant memory codes may be most frequent in tasks which place a premium on variability of encoding and interrelating of stimuli. Where task requirements are more limited, so that only a single code for each of a limited number of items must be held in STS, there may be only a sporadic loosening on the restrictions on information flow from LTS. Thus, in free recall learning where optimal performance is achieved only when subjects utilize alternative encodings of items and associate items with one another, there may be a continual flow of irrelevant codes for list items and their non-list associates. On the other hand, in psychomotor tasks, such as simple or choice reaction time, the repetitive nature of the task may allow intoxicated subjects to restrict the flow of irrelevant information into STS during most
trials of the task, with performance dropping significantly only on those trials where the restrictions are loosened. This could explain why some investigators have shown that cannabis effects on psychomotor tasks seem to be intermittent, as if subjects were distracted on a portion of the trials (Clark et al., 1970).
This model makes many of the same predictions and is conceptually similar to a model proposed by DeLong and Levy (1973). They suggested that cannabis-induced behavioral changes could be understood in terms of various aspects of attention including focusing on relevant aspects of a task and resisting distraction. In the model presented here, the influx of irrelevant material from LTS serves, in a sense, as distracting input which inhibits the processing of task-relevant information. A major contribution of the present model is that it not only explains measurable behavioral effects of cannabis but also reported subjective effects of the drug.
In many memory models, the flow of information between memory stores, retrieval of stored information, complex coding operations and many other higher cognitive functions are under the control of an 'executive monitor' or 'executive program'. To meet task requirements the executive monitor must control the flow of memory codes from LTS into STS. Thus, the monitor may be the focus of cannabis effects since such a flow of information seems to be altered by the drug. One direction of cannabis research may be to examine drug effects on other activities of this monitor. Experiments by Hart (1965) have demonstrated a process he calls 'memory monitoring' which allows individuals to examine their memories to determine if currently irretrievable information is potentially available for recall. Such a process enables the individual to decide whether, after a series of retrieval failures, further similar attempts should be made, retrieval strategies should be altered, or retrieval effort should be abandoned altogether. By utilizing variants of Hart's paradigm, future research may delineate effects of cannabis on the processes which control and direct subjects' behavior and which may be ultimately responsible for the varied psychological and cognitive effects induced by the drug.
Future research in the psychological and cognitive effects of cannabis might also usefully include various pharmacological manipulations. One such line of investigation entails the use of other drugs to either inhibit specific effects of cannabis or to interact with the behavioural characteristics of cannabis in other ways. For example, although it has been established that there is little or no cross tolerance with LSD, knowledge about cross tolerance with other drugs is limited (Isbell and Jasinski, 1969). Information about such interactions sometimes provides clues about which receptor sites or physiological systems are responsible for certain behavioural effects of the drug. Similarly information about which agents block drug effects could be exceedingly useful. Except for studies indicating that the cognitive effects of cannabis are not significantly reduced by pre-treatment with the beta-adrenergic blocker, propranolol, little work has been completed in this area ( Drew et al., 1972). Pharmacological differences among various batches of cannabis may also provide useful information. For example, although most of the effects of acute intoxication with cannabis can be explained on the basis of delta-l-THC (delta-9-THC by other nomenclature) and its metabolites, it is possible that other components of cannabis or their metabolites selectively exert certain behavioural effects. Cannabis would thus be directly comparable to alcoholic beverages in which the predominant effects are exerted by ethanol, but various congeners 9ay also induce discernible influences (Katkin et al., 1970). Such a finding might also explain reports suggesting that various cannabis preparations exert different psychological effects.
A final area of psychological and cognitive research with cannabis that seems promising involves comparing the psychological and cognitive effects of cannabis with other psychoactive agents and hormones (Drew and Miller, 1974). Cannabis seems to occupy a unique position in the pharmacopoeia; some of its properties are analogous to the sedative hypnotics such as alcohol and secobarbital, but other effects are comparable to low doses of LSD and other psychedelic substances. Comparing the effects of cannabis with other drugs is useful not only at a psychological and cognitive level, but also in attempts to determine the physiological mechanisms that subserve these behavioral effects. Integrating the psychological and cognitive effects of cannabis with the underlying alterations in physiological processes remains one of the ultimate goals of cannabis research.
REFERENCES
Abel, E.L. Marihuana and memory: acquisition or retrieval? Science, 173: 1038-1040, 1971.
Ames, F. A clinical and metabolic study of acute intoxication with cannabis sativa and its role in a model psychosis. Journal of Mental Science, 104: 972-999, 1958.
Anisfield, M., and Knapp, M. Association, synonymity and directionality in false recognition. Journal of Experimental Psychology, 77: 171-179, 1968.
Atkinson, R.C. and S hiffrin, R.M. Human memory: a proposed system and its control processes. In K.W. Spence and J.T. Spence, eds. The Psychology of Learning and Motivation. Vol. 11, pp.89-195. Academic Press, New York, 1968.
Atkinson, R.C. and Shiffrin, R.M. The control of short-term memory. Scientific American, 224: 82-90, 1971.
Atkinson, R.C., Herrmann, D.J. and Wescourt, K.T. Search Processes in Recognition Memory. Technical Report No. 204, Institute for Mathematical Studies in the Social Sciences, Stanford University, June 8, 1973.
Bousfield, W .A., Whitmarsh, G.A. and Danick, J.J. Partial response identities in verbal generalization. Psychological Reports, 4: 703-713, 1958.
Bromberg, W. Marihuana intoxication, a clinical study of cannabis sativa intoxication. American Journal of Psychiatry, 91: 303-330, 1934.
Caldwell, D.F., Myers, S.A., Domino, E.F. and Merriam, P.E. Auditory and visual threshold effects of marihuana in man. Perceptual and Motor Skills, 29: 755-759, 1969.
Caldwell, D.F., Myers, S.A. and Domino, E.F. Effects of marihuana on sensory thresholds in man. In D.H. Efron, ed. Psychotomimetic Drugs. pp.299-321. Raven Press, New York, 1970. Canada. Commission of Inquiry into the Non-Medical Use of Drugs. Cannabis. Queen's Printer, Ottawa, 1972.
Cappel, H., W ebster, C.E., Herring, B.S. and Ginsberg, R. Alcohol and marihuana: a comparison of effects on a temporally controlled operant in humans. The Journal of Pharmacology and Experimental Therapeutics, 182 (2): 195-203, 1972.
Casswell, S. and Marks, D.F. Cannabis and temporal disintegration in experienced and naive subjects. Science, 179: 803-805, 1973.
Chopra, R.N. and Chopra, G.S. The present position of hemp-drug addiction in India. Indian Journal of Medical Research Memoirs, 31: 1-119, 1939.
Clark, L.D. and Nakashima, E.H. Experimental studies of marihuana. American Journal of Psychiatry, 125: 379-384, 1968.
Clark, L.D., Hughes, R. and Nakashima, E.N. Behavioral effects of marihuana: experimental studies. Archives of General Psychiatry, 23: 193-197, 1970.
Craik, F.I.M. The fate of primary memory time in free recall. Journal of Verbal Learning and Verbal Behavior, 9: 672-678, 1970.
Darley, C.F., Tinklenberg, J.R., Hollister, L.E. and Atkinson, R.C. Marihuana and retrieval from short-term memory. Psychopharmacologia (Berl.), 29: 231-238, 1973a.
Darley, C.F., Tinklenberg, J.R., Roth, W.T., Hollister, L.E. and Atkinson, R.C. Influence of marihuana on storage and retrieval processes in memory. Memory and Cognition, 1: 196-200, 1973b.
Darley, C.F. and Tinklenberg, J.R. Marihuana and memory. In L.L. Miller and W.G. Drew, eds. Marijuana - Effects of Human Behavior. Academic Press, New York, in press.
Darley, C.F., Tinklenberg, J.R., Roth, W.T. and Atkinson, R.C. The nature of storage deficits and state-dependent retrieval under marihuana. Psychopharmacologia (Berl.), in press.
DeLong, F.L. and Levy, B.I. Cognitive effects of marijuana, described in terms of a model of attention. Psychological Reports, 33: 907-916, 1973.
Dornbush, R.L. Marijuana and memory: effects of smoking on storage. Transactions of the New York Academy of Sciences, 36: 94-101, 1974.
Dornbush, R.L., Fink, M. and Freedman, A.M. Marihuana, memory and perception. American Journal of Psychiatry, 128: 194-197, 1971.
Drew, W.G. and Miller, L.L. Cannabis: neural mechanisms and behavior - a theoretical review. Pharmacology, 11 (1): 12-33, 1974.
Drew, W.G., Kiplinger, G.F., Miller, L.L. and Marx, M. Effects of propranlol on marihuana-induced cognitive dysfunctioning. Clinical Pharmacology and Therapeutics, 13 (4): 526-533, 1972.
Eich, J., Weingartner, H. and Stillman, R.C. Availability versus accessibility of information in marijuana state-dependent learning. Paper submitted for publication, 1973.
Galanter, M., Wyatt, R.J., Lamberger, L., Weingartner, H., Vaughn, T.B. and Roth, W.T. Effects on humans of delta-9-tetrahydrocannabinol administered by smoking. Science, 176: 934-936, 1972.
Glanzer, M. and Cuntiz, A.R. Two storage mechanisms in free recall. Journal of Verbal Learning and Verbal Behavior, 5: 351-360, 1966.
Hart, J.T. Recall, recognition and the memory-monitoring process. Unpublished Ph.D. Thesis, Stanford University, 1965.
Hochman, J.L. and Brill, N.Q. Marijuana use and psycho-social adaptation. Paper delivered at American Psychiatric Association meeting in Washington, D.C., May 3, 1971.
Hollister, L.E. Marihuana in man: three years later. Science, 172: 21-28, 1971.
Hollister, L.E. and Gillespie, H.K. Marihuana, ethanol and dextroamphetamine. Mood and mental function alterations. Archives of General Psychiatry, 23: 199-203, 1970.
Hollister, L.E. and Tinklenberg, J.R. Subchronic oral doses of marihuana extract. Psychopharmacologia (Berl.), 29: 247-252, 1973.
Hollister, L.E. Department of Medicine and Psychiatry, VA Hospital, Palo Alto and Stanford University. Personal communication, April, 1974.
Isbell, H. and Jasinski, D.R. A comparison of LSD-25 with (-)-delta-9-transtetrahydrocannabinol (TUC) and attempted cross tolerance between LSD and THC. Psychopharmacologia (Berl.), 14 (2): 115-123, 1969.
Isbell, H., Gorodetzsky, C.W., Jasinski, D., Claussen, U., Spulak, F.D. and Korte, F. Effects of (-)delta-9-trans-tetrahydrocannabinol in man. Psychopharmacologia (Berl.), 11: 184-188, 1967.
Jones, R.T. Tetrahydrocannibinol and the marijuana induced social 'high' or the effects of the mind on marijuana. Annals of the New York Academy of Sciences, Vol. 191. A.J. Singer, ed. New York Academy of Sciences, New York, 1971.
Jones, R.T. Department of Psychiatry, Langley Porter Clinic, University of California. Personal communication, April, 1974.
Jones, R.T. and Stone, G.C. Psychological studies of marijuana and alcohol in man. Psychopharmacologia (Berl.), 18: 108-117, 1970.
Karniol, I.G. and Carlini, E.A. Comparative studies in man and in laboratory animals on delta-8- and delta-9-trans-tetrahydrocannabinol. Pharmacology, 9: 115-126, 1973.
Katkin, E.S., Hayes, W.N., Teger, A.I. and Pruitt, D.E. Effects of alcoholic beverages differing in congener content on psychomotor tasks and risk-taking. Quarterly Journal of Studies on Alcohol, Supplement, 5: 101-114, 1970.
Kiplinger, G.F., Manno, J.E., Rodda, B.E. and Forney, R.B. Dose-response analysis of the effects of tetrahydrocannabinol in man. Clinical Pharmacology and Therapeutics, 12: 650-657, 1971.
Klonoff, H„ Low, M. and Marcus, A. Neuropsychological effects of marijuana. Canadian Medical Association Journal, 108: 150-157, 1973.
Lemberger, C.M., Silberstein, S.D., Axelrod, J. and Kopin, I.J. Marihuana: studies on the disposition and metabolism of delta-9-tetrahydrocannabinol in man. Science, 170: 1320-1322, 1970.
Manno, J.E., Kiplinger, G.F., Haine, S.E., Bennett, I.F. and Forney, R.B. Comparative effects of smoking marihuana or placebo on human motor and mental performance. Clinical Pharmacology and Therapeutics, 11: 808-815, 1970.
Manno, Kiplinger, T.F., Scholz, N., and Forney, R.B. The influence of alcohol and marihuana on motor and mental performance. Clinical Pharmacology and Therapeutics, 12: 202-211, 1971.
Melges, F.T., Tinklenberg, J.R., Hollister, L.E. and Gillespie, H.K. Temporal disintegration and depersonalization during marihuana intoxication. Archives of General Psychiatry, 23: 204-210, 1970a.
Melges, F.T., Tinklenberg, J.R., Hollister, L.E. and Gillespie, H.K. Marihuana and temporal disintegration. Science, 168: 1118-1120, 1970b.
Melges, F.T., Tinklenberg, J.R., Deardorff, C.M., Davies, N.H.,, Anderson, R.C. and Owen, C.A. Temporal disorganization and delusional-like ideation. Archives of General Psychiatry, in press.
Melton, A.W . and Martin, E. Coding Processes in Human Memory. V.H. Winston and Sons, Washington, D.C., 1972.
Mendelson, J.H. and Meyer, R.E. Behavioral and biological concomitants of chronic marihuana smoking by heavy and casual users. In United States, National Commission on Marihuana and Drug Abuse, Marihuana: A Signal of
Misunderstanding, Appendix, Vol. 1, pp. 68-246. U.S. Government Printing Office, Washington, D.C., 1972.
Meyer, R.E., Pillard, R.C., Sapiro, L.M. and Mirin, S.M. Administration of marihuana to heavy and casual marihuana users. American Journal of Psychiatry, 128: 198-204, 1971.
Miller, L.L., Drew, W.G. and Kiplinger, G.F. Effects of marijuana on recall of narrative material and Stroop colour-word performance. Nature (Lond.), 237: 172-173, 1972.
Moskow itz, H., Sharma, S. and Schapero, M. A comparison of the effects of marijuana and alcohol on visual functions. In M.F. Lewis, ed. Current Research in Marihuana, pp.129-150 . Academic Press, New York, 1972.
Norman, D.A., ed. Models of Human Memory. Academic Press, New York, 1970.
Norman, D.A. and Rumelhart, D.E. A system for perception and memory. In D.A. Norman, ed. Models of Human Memory, pp.19-64. Academic Press, New York, 1970.
Ornstein, R.E. On the Experience of Time. Penguin, Baltimore, 1969.
Pearl, J.H., Domino, E.F. and Rennick, P. Short-term effects of marijuana smoking on cognitive behavior in experienced male users. Psychopharmacologia (Berl.), 31: 13-24, 1973.
Perez-Reyes, M., Lipton, M.A., Timmons, M.C., Wall, M.E., Brine, D.R. and Davis, K.H. Pharmacology of orally administered delta-9-tetrahydrocannabinol. Clinical Pharmacology and Therapeutics, 14: 48-55, 1973.
Rafaelsen, 0.J., Bech, P., Christiansen, J., Christrup, H., Nyboe, J. and Rafaelsen, L. Cannabis and alcohol: effects on simulated car driving. Science, 179: 920- 923, 19 73.
Renault, P.F., Schuster, C.R., Heinrich, R. and Fredman, D.X. Marijuana: standardized smoke administration and dose effect curves on heart rate in humans. Science, 174: 589-591, 1971.
Rickles, W .H., Jr., Cohen, M.J., Whitaker, C.A. and McIntyre, K.E. Marijuana induced state-dependent verbal learning. Psychopharmacologia (Berl.), 30: 349-354, 1973.
Roth, W.T., Galanter, M., Weingartner, H., Vaughan, T.B. and Wyatt, R.J. Marijuana and synthetic delta-9-THC: some effects on the auditory evoked response and background EEG in humans. Biological Psychiatry, 6: 221-233, 1973.
Sperling, Go The information available in brief visual presentations. Psychology Monographs, whole no. 498, 1960.
Tart, C.T. Marihuana intoxication: common experiences. Nature, 226: 701-704, 1970.
Tinklenberg, J.R., Melges, F.T., Hollister, L.E. and Gillespie, H.K. Marijuana and immediate memory. Nature, 226: 1171-1172, 1970.
Tinklenberg, J.R., Kopell, B.S., Melges, F.T. and Hollister, L.E. Marihuana and alcohol: time production and memory functions. Archives of General Psychiatry, 27: 812-815, 1972.
Tulving, E. and Donaldson, W.D., eds. Organization of Memory. Academic Press, New York, 1972.
Underwood, B.J. False recognition produced by implicit verbal responses. Journal of Experimental Psychology, 70: 122- 129, 1965.
Underwood, B.J. Attributes of memory. Psychological Review, 76: 559-573, 1969.
Volavka, J., Dornbush, R., Feldstein, S., Clare, G., Zaks, A., Fink, M. and Freedman, A.M. Marijuana, EEG, and behavior. Annals of the New York Academy of Sciences, Vol. 191. A.J. Singer, ed. New York Academy of Sciences, New York, 1971.
Weil, A.T. and Zinberg, N.E. Acute effects of marihuana on speech. Nature, 222: 434-437, 1969.
Weil, A.T., Zinberg, N.E. and Nelson, J.M. Clinical and psychological effects of marijuana in man. Science, 162: 1234-1242, 1968.
DISCUSSION
As in all discussions at this conference the subject matter of the paper was discussed at length and in depth. In order to organise the material, subject headings have been chosen and relevant contributions placed within these headings rather than sticking strictly to the consecutive order of the contributions. The resulting statement is a summary overview of main themes of the discussion.
SUBJECTS AND DOSAGE LEVELS
In response to a question by Dr Rubin, Dr Tinklenberg described the choice of subjects for the study. He noted that it had not been possible to use naive subjects because of the possible danger to corrupting them, nor indeed to use women. He therefore had no data on naive users. Experience had shown that individuals over 30 years of age such as school superintendents, police, court judges and others were not suitable because they were more likely
to react badly to cannabis. His subjects, therefore, were non-naive graduate students age 19-24 who had used 'occasionally' or up to once or twice a week for the last year. Previous experience had shown that it was advisable to avoid medical students, who tended to be neurotic, and psychology or behavioral students who tended to manipulate the study . Stanford University graduate students of civil engineering were found to be particularly suitable since they were reliable, bright, compulsive and obedient.
In response to a question by Dr Martens, Dr Tinklenberg stated that the dose used in the study was 20 mg. THC orally, equivalent to a smoking dose of 6-7 mg. per person and described by the subjects as a fairly strong dose. Material was supplied by NIMH and dosage levels could therefore be compared with other studies using NIMH material.
THE MODEL PRESENTED
It is useful at this point to summarise the main features of the model of sensory processing put forward by Dr Tinklenberg. The model postulates that sensory input is brought into relationship with a conceptual store of previously encoded inputs, and an encoded representation of the input proceeds to short-term memory, where it becomes conscious and open to control processes, such as rehearsal and imagery. Rehearsal may then place the encoded input in the Event Knowledge Store of Long-Term Memory.
Dr Tinklenberg's suggestion is that, under cannabis intoxication, a greater number of codes than usual is activated by sensory inputs. Because short-term memory can only handle a limited number of codes at a time, the arrival of a greater number of codes causes previously encoded stimuli to be dropped more quickly than otherwise from short-term memory. Transfer to long-term memory is therefore impaired. The model also offers explanations of increase of intrusion errors, acceleration of subjective time-sense, subjective increase in richness of perception, and formation of new 'creative' associations.
THE CODING PROCESS
Dr Mortens asked whether the model merely postulated a greater number of activated codes under intoxication, or whether a different type of code might be activated. The significance of the distinction, he suggested, is that a change in choice of codes would imply a more central effect than would a simple increase in number of codes activated. Dr Tinklenberg replied that existing evidence pointed to a greater number rather than a different choice of codes, and that an experiment was planned to investigate this.
Dr Edwards raised the question of a possible analogy between the greater number of codes activated by cannabis intoxication and the kind of overinclusive thinking sometimes observed in schizophrenia. Dr Tinklenberg suggested that alterations in the schizophrenic's processing of incoming information occurred at an earlier stage of processing than was the case with cannabis intoxication.
ATTENTION
Dr Smart asked about the power of the model in explaining results of studies of attention. Although sensory registration studies (undivided attention, single stimulus) may not show a decrease in performance under cannabis intoxication, studies of divided attention do show deficits. Divided attention studies are more like everyday life.
Dr Tinklenberg agreed that such results are not explained parsimoniously by the model, but suggested that whatever was happening was at a central rather than a peripheral level. Professor Paton suggested that the addition of a feedback loop allowing central direction of input might be helpful.
Dr Leuw raised the question of motivation as an intervening variable and noted that motivation could overcome the effects of cannabis. Dr Tinklenberg agreed but stated that on certain serial learning tasks there was no significant difference between those under the influence of cannabis and those not under its influence.
Dr Hindmarch mentioned task-dependent performance, and stressed that performance could be drastically altered by differences in how one directed the subject. He felt that the effects of motivational and task-dependent factors could be unravelled by careful experimental design.
Whilst a number of participants found the model proposed to be interesting and a valuable way of considering a difficult area of cognitive function several participants, notably Drs Hindmarch, Leuw and Smart, were concerned that the model was a little mechanistic and was abstracted memory from other systems. As Dr Smart put it:
'I'm always very happy to see theories develop and models presented. I rarely attempt them myself. This one still bothers me - especially in respect of motivation. There are studies that have shown that deficits under cannabis can be experimentally manipulated. We need to add another box to the model for unknowables.'
Acknowledgements:
The authors wish to thank Patricia L. Murphy and Peggy Murphy for special assistance in the preparation of this paper as well as Cynthia Demos for technical assistance.
The authors' research discussed in this paper was supported in part by the van Ameringen Foundation.
| < Prev | Next > |
|---|